Chao Zhao1,2,
Chao Ai1,2,
Qiuzhe Li1,3,
Chengfeng Yang1,3,
Guangzhi Zhou1,3,
Bin Liu1,3,4 ![]()
For correspondence:-
Received: 10 September 2015 Accepted: 20 November 2016 Published: 21 December 2016
Citation: Zhao C, Ai C, Li Q, Yang C, Zhou G, Liu B. Diversity of archaea and bacteria in a biogas reactor fed with Pennisetum sinese Roxb by 16S rRNA sequence analysis. Trop J Pharm Res 2016; 15(12):2659-2667 doi: 10.4314/tjpr.v15i12.18
© 2016 The authors.
This is an Open Access article that uses a funding model which does not charge readers or their institutions for access and distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/4.0) and the Budapest Open Access Initiative (http://www.budapestopenaccessinitiative.org/read), which permit unrestricted use, distribution, and reproduction in any medium, provided the original work is properly credited..
Purpose: To investigate the structure and function of the complex rumen microbial community in a biogas reactor by 16S rRNA gene analysis, which was fed with Pennisetum sinese Roxb as the mono-substrate.
Methods: Two 16S ribosomal RNA (rRNA) clone libraries of bacteria and archaea were established by polymerase chain reaction. Community structure was determined by phylogenetic analyses of 119 and 100 16S rRNA gene clones from the bacterial and archaeal libraries, respectively.
Results: In the bacterial library, 13.4 % of clones were affiliated with Treponema porcinum, 5.9 % with Eubacterium limosum, 5 % with Clostridium, 5 % with Bacteroidetes, 4.2 % with Firmicutes, 2.5 % with Anaerofilum and a total of 64 % clones belonged to unclassified or uncultured bacteria. In the archaeal library, Methanobacterium curvum made up 12 % of known clones, Methanosarcina barkeri represented 8 %, Methanobacterium bryantii represented 4 % and Methanofollis ethanolicus represented 2 %, respectively; the remaining 74 % of the clones were unclassified archaebacteria.
Conclusion: T. porcinum and M. curvum are the most predominant bacteria and archaea in a biogas reactor fed with P. sinese as the sole substrate.
Introduction
Biogas is a highly relevant form of bioenergy for communities. A wide variety of renewable biomass feedstocks can be used for energy, including agricultural residues and energy crops. Large numbers of acid- and methane-producing anaerobic bacteria convert organic matter into methane gas in an anaerobic digestion process [1].
Pennisetum sinese Roxb (PSR), a species of fast-growing grass common to large parts of the world is used in biogas systems [2]. PSR has a high energy-conversion efficiency; its solar energy conversion rate is four- to six-fold greater than that of broad-leaved trees. Its yields are five- to ten-fold than those of the crops and reaches 450 tons per hectare. Due to its favourable properties, it has potential as an industrial feedstock for biogas production.
The rumen is a complex habitat in which feedstuffs are fermented primarily into a mixture of volatile fatty acids, which serve as a major source of nutrients for ruminant animals. Most of rumen micro-organisms are unculturable and our knowledge of the microbial diversity is thus incomplete. Several recent studies have identified the most abundant species in the rumen [3]. Recently, the potential applications of rumen microbes in artificial reactors for the conversion of cellulose-rich materials have been explored [4].
Bioconversion of PSR can provide energy and reduce deforestation in industrial applications. However, little work has been reported on rumen microbial fermentation with PSR. In this study, we investigated the structure and function of the complex rumen microbial community in a biogas reactor by 16S rRNA gene analysis, which was fed with PSR as the sole substrate.
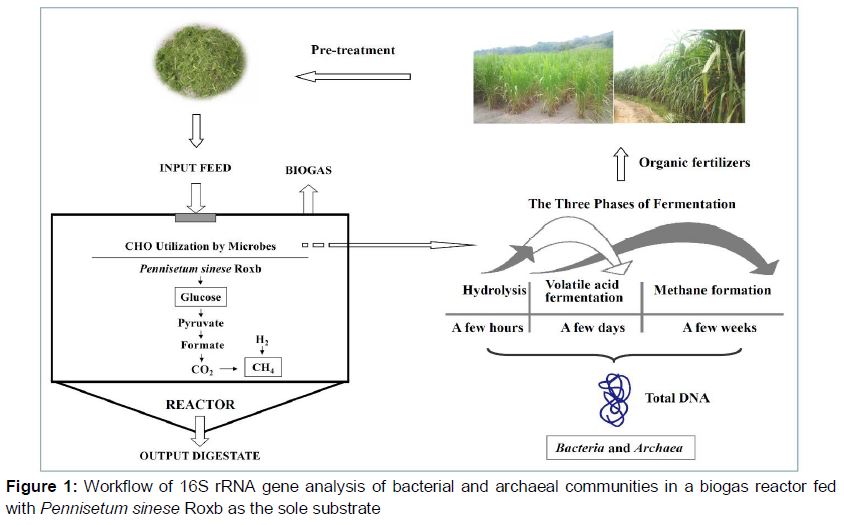
The process of workflows () can be described as follows: The harvested PSR was crashed to granules by fodder grinder and put into the anaerobic fermentation reactor for 15 days; then 16s rRNA library was constructed to reflect the potential information of microbial fermentation community.
Methods
Rumen sampling, biogas reactor description and sampling
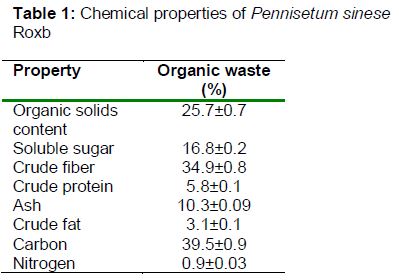
Cows were maintained in adjacent indoor tie stalls according to a protocol approved by the College of Animal Science, Fujian Agriculture and Forestry University (FAFU). They were allowed ad libitum access to feed and water, and enough feed was supplied once daily as a total mixed ration. The scientific procedure was used to feed animal based on the Guide for the Care and Use of Laboratory Animals [5]. The ethical review board of Fuzhou General Hospital of Nanjing Military Command provided ethical approval (number FZJQ2011018). The faeces samples were collected and stored at -80 °C until further analysis. Fresh PSR material was identified and provided by Prof. Zhanxi Lin of the China National Engineering Research Center of JUNCAO Technology (Fuzhou, China). The growth period was 90 days. Dry matter, organic solids content, soluble sugar, crude fibre, crude protein, ash, crude fat, carbon, and nitrogen contents were measured by the Institute of Bioenergy, FAFU. The chemical properties required for the production of biogas are shown in the . The carbon-nitrogen (C/N) ratio of PSR was 42.5, which was suitable for producing biogas. An anaerobic fermentation tank (5 L) with a mixer was used as the biogas reactor, and was supplied with 0.5 kg PSR.
The total gas volume and methane content of the biogas produced were 14.5 L and 6.2 L, respectively. The biogas reactor was run at high temperature (55 °C) under wet fermentation conditions at approximately pH 6.8. The retention time for continuous fermentation was 15 days. Two litres of completely fermented slurry sample, which could be treated as a steady-state sample with a constant community structure, were collected from the bottom of the fermentation tank by pumping.
Collection of community DNA
The bacterial and archaeal community members in this biogas reactor fed with PSR were investigated. Samples (50 mL) of slurry were centrifuged at 12,000 g for 5 min, and the supernatant was decanted. Bacterial cells were collected on day 15 of culture, during the fermentation period. Genomic DNA was extracted using a Fast DNA® Spin Kit for Soil (Mobio, USA) and according to the manufacturer’s instructions.
PCR amplification and clone library construction
Total genomic DNA from anaerobic digestion sludge was used as a template to construct 16S rRNA libraries of the bacterial and archaeal communities. PCR amplification was carried out in 50 μL reaction volumes, containing 100 ng genomic DNA, 10 × reaction buffers, 20 pmoL of each primer, 0.4 mM (each) dNTP, and 1.0 U rTaq DNA polymerase (TaKaRa). Briefly, the bacterial 16S rRNA gene fragment was amplified using the universal bacterial primers 27F and 1492R. The following PCR program was used to amplify the full-length bacterial 16S rRNA gene: initial denaturation for 5 min at 94 °C; 30 cycles of 94 °C for 45 s, 58 °C for 45 s, and 72 °C for 1.5 min; and a final elongation for 10 min at 72 °C. To amplify the archaeal 16S rRNA genes, the archaea-specific primers 21F-958R and the protocol of Kolganova et al were used [5]. The PCR procedure included an initial cycle of 5 min at 94 °C, followed by 20 cycles of 1 min at 94 °C, 45 s at 55 °C, and 1 min at 72 °C, with a touchdown of 0.5 °C per cycle, followed by 16 cycles of 1 min at 94 °C, 45 s at 50 °C, and 1 min at 72 °C; for the final extension, the reaction was held at 72 °C for 10 min. The purified PCR product was cloned into pMD18-T, transformed into Escherichia coli DH5α competent cells (TaKaRa) as per the manufacturer’s protocol, and sequenced using the M13 primers by Invitrogen Biotechnology.
Phylogenetic analysis and tree construction of bacteria and archaea species
16S rRNA libraries of the microbial community were constructed. The PCR-amplified full-length 16S rRNA gene fragments were cloned into the pMD18-T vector (TaKaRa). More than 100 clones were randomly selected and checked for the correct insert size by agarose gel electrophoresis and sequencing. The sequences of 16S rRNA genes were aligned with sequences from the GenBank database using the NCBI BLASTN comparison software.
Statistical analysis
Phylogenetic trees were constructed using neighbour-joining method with the MEGA software (version 6.0) with 1,000 bootstrap replicates. The 16S rRNA gene sequences of the most highly phylogenetically related strains were selected as reference strains. Community strains and selected reference strains were evaluated phylogenetically by nucleotide sequencing of the 16S rDNA.
Results
Bacterial community in biogas reactor
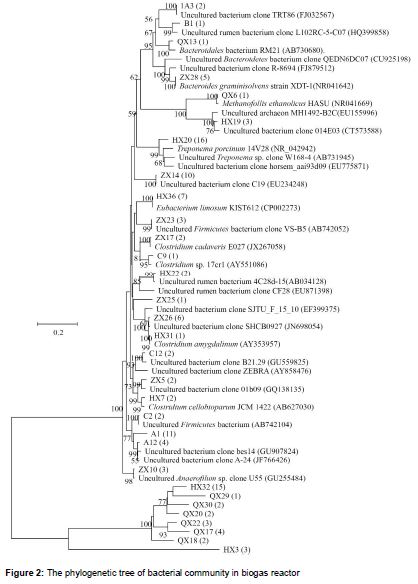
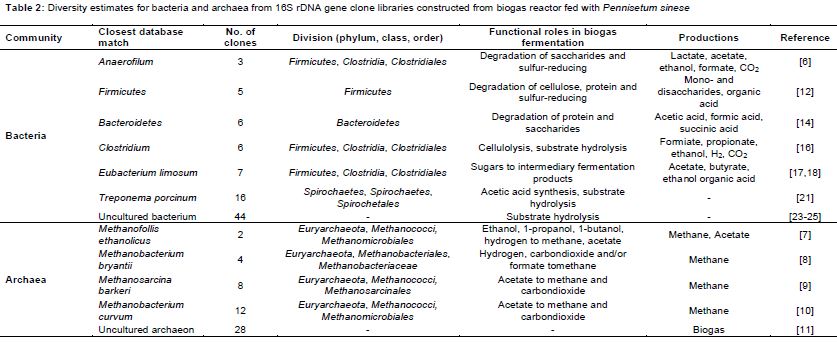
A total of 219 clones were detected and sequenced, which consisted of 119 clones of bacteria and 100 clones of archaea. Statistical analysis of 16S rRNA gene libraries indicated that all major groups of bacteria and archaea from the biogas reactor had been detected. Examination of 43 clones showed that, with the phylogenetic diversity, bacterial community was covered most of the rumen bacterial species in the library while were fed with PSR as the sole substrate (). The sequenced clones fell into the following six major lineages: Anaerofilum, Firmicutes, Bacteroidetes, Clostridium, Eubacterium, Treponema, and related micro-organisms. Of the clones, 2.5 % belonged to Anaerofilum, which may originate from the chicken intestine and under anaerobic conditions, produces hydrogen. A further 4.2 % of the clones were Firmicutes, which made up 30.1 % of the sequences in the clone library.
Archaeal community in the biogas reactor
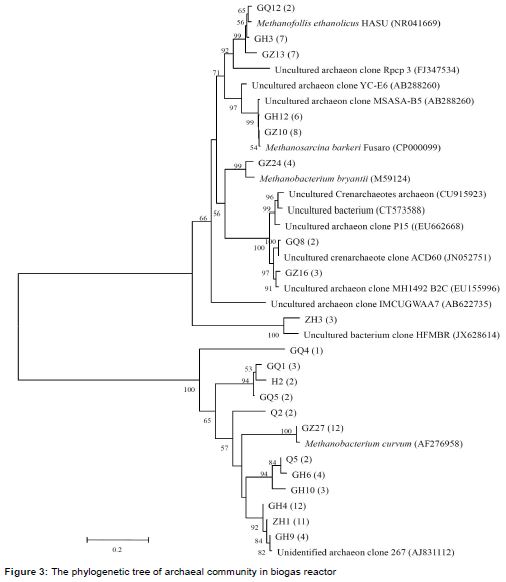
A 16S rRNA gene clone library of archaeal community was also constructed, and 100 clones were sequenced (). Based on the alignment and phylogenetic analyses of these sequences, 26 % of the clones were classified into four archaeal taxonomic groups (). Methanofollis ethanolicus, which converted ethanol, 1-propanol, 1-butanol, and hydrogen into methane and acetate, accounted for 2 % [7].
Methanobacterium bryantii represented 4 %, and are capable of converting hydrogen and/or formate to CH4 [8]. Methanosarcina barkeri (8 %), an acetoclastic methanogen, produced CH4 from methyl compounds such as methanol and methylamines [9]. The remaining 12 % were accounted for by Methanobacterium curvum, which can utilize acetate for methane production via the hydrogenotrophic and methylotrophic pathways [10]. Furthermore, 28 % the clones were uncultured archaeon, which are important for the biodegradation of organic compounds and might play a vital role in methane production [11]. The remaining 46 % were considered unclassified clones and were predicted to be involved in biogas production.
Culturable bacterial species represent only a small fraction of the total bacterial diversity. The rumen is no exception. The features and functions of rumen bacteria are listed in . The readily culturable bacterial population of the rumen (as estimated by viable cell count in rich media) made up 5–15 % of the total bacterial population (determined by direct cell count).
Discussion
Members of the phylum Firmicutes are the major degraders of cellulose, although few express glycoside hydrolases [12]. This phylum represents 32 % of the bacterial community in wet fermentation conditions and 25 % in dry fermentation conditions, respectively [13]. Furthermore, 5 % of sequences were identified as Bacteroidetes, which are involved in the degradation of complex polysaccharides [14]. It is the second-most abundant bacterial taxon in lignocellulose degradation cultures, and the third-most abundant bacteria in the gut of the lower termite Coptotermes [15]. Clostridium, which accounted for 5.0 % of the clones, is an anaerobic cellulolytic bacterium [16]. Eubacterium limosum, which represented 5.9 %, is a strictly anaerobic gram-positive bacterium with demethylation activity that is part of the human intestinal microflora [17]. Song and Cho reported that E. limosum ATCC 8486 converted and transformed syngas and isoflavonoids into butanol and phytoestrogens, respectively [18]. An anaerobic syngas-converting bacteria, the genome of E. limosum KIST612, harbours genes encoding the key enzymes for conversion of acetyl-CoA into bioenergy-compatible acids/alcohols (acetate, butyrate, and ethanol) [19]. T. porcinum, a homoacetogenic genus that is enriched with the addition of biomethanation of coke oven gas, represented the remaining 13.4 % of the clones [20]. A study of anaerobic co-fermentation of straw and manure suggested that T. porcinum is specialized to produce acetate by utilizing H2 and CO2 [21].
Forty-four clones of uncultured rumen bacteria were identified. With the development of DNA sequencing technology, more and more functional genes have been found in uncultured rumen bacterium. Cellulose-degrading microbes are commonly adherent to plant cell walls. A lack of understanding of how to exploit this process may be one of the reasons that limited the inoculation of rumen microorganisms [22]. Novel xylanase genes have been cloned from uncultured strains in rumen fluid for use as a feed additive due to the high digestibility of the rumen toward cellulosic stuffs [23]. Besides, a novel uncultured environmental strain, 7WAY-8-7(T), is capable of converting glucose into acetate and hydrogen [24]. The considerable ecological significance of uncultured bacteria in the rumen is supported by the positive interaction between strains R-25 and F. Their activities of carboxy-methylcellulose and xylanase can be increased by further co-culture [25]. However, the remaining 32 clones were unclassified bacteria; therefore, as-yet-unknown species may be involved in substrate degradation.
Rumen methanogens are classified into three groups according to their metabolism: hydrogenotrophs, methylotrophs, and acetoclastic methanogens. Hydrogenotrophs (Methanobrevibacter, Methanomicrobium, and Methanobacterium) convert hydrogen and/or formate into CH4, while methylotrophs (Methanosphaera and members of the order Methanomassiliicoccales) produce CH4 from methyl compounds such as methanol and methylamines. Acetoclastic methanogens (Methanosarcina) use acetate to produce CH4 through the pathway that in addition to the hydrogenotrophic and methylotrophic pathways. Methanogenes, which were discovered in the biomass of anoxic biogas reactors, was catalysed by the syntrophic cooperation of anaerobic bacteria, syntrophic acetogenic bacteria, and methanogenic archaea. Analysis of microbial community composition may improve biogas yields from biomass. Compared to previous reports that hydrogenotrophic methanogens are the dominant species in biogas plants, our results indicate that M. curvum was the predominant methanogenic archaeal species in a biogas reactor with PSR as the sole substrate.
Conclusion
T. porcinum and M. curvum are the most predominant bacteria and archaea in the biogas reactor. In further studies, metagenomics and metatranscriptomics will be used to analyse the functional relationships among members of microbial biogas communities.
Declarations
Acknowledgement
References
Archives


News Updates